TIPOS DE COENZIMAS Y VITAMINAS
Se describirán las principales
vitaminas que se conocen en la actualidad la naturaleza de sus efectos
biológicos, los coenzimas de los cuales las vitaminas son componentes
esenciales, y se darán ejemplos de la función coenzimas en las reacciones
metabólicos específicas catalizadas por los enzimas
TABLA. Vitaminas y sus formas coenzimaticas.
|
TIPO
|
COENZIMA O FORMA ACTIVA
|
FUNCION PROMOVIDA
|
|
HIDROSOLUBLES
|
|
|
|
Tiamina
Riboflavina
Acido nicotínico
Ácido pantotecnico
Piridoxina
Biotina
Acido fólico
Vitamina B12
Ácido lipoico
Ácido ascórbico
|
Pirofosfato
de tiamina (TPP)
Flavin-mononucleotido(FMN)
Flavin-adenina
-dinucleótido(FAD)
Dinucleotido
de nicotidamina y de adenina(NAD)
Dinucleotido
de nicotidamina y de adenina fosfato (NADP)
Coenzima
A (CoA)
Fosfato
de piridoxal
Biocitina
Ácido
tetrahidrofolico
Coenzima
B12
Lipoil-lisina
--------------
|
Transferencia de grupo aldehído
Transferencia
de átomo de hidrogeno (electrón )
Transferencia
de átomo de hidrogeno (electrón )
Transferencia
de átomo de hidrogeno (electrón )
Dinucleotido
de nicotidamina y de adenina fosfato (NAD)
Transferencia
de grupo acilo
Transferencia
de grupo amino
Transferencia
de carboxilo
Transferencia
de grupo monocarbon
Desplazamiento
de 1,2 átomos de hidrógeno y de grupo acilo
Cofactor
de hidroxilación
|
|
LIPOSOLUBLES
|
|
|
|
Vitamina A
Vitamina D
Vitamina E
Vitamina K
|
11-cis-retinal
1,25-dihidroxicolecalciferol
-----
-----
|
Ciclo
visual
metabolismo
de calcio y del fosfato
antioxidante
biosíntesis
de protrombina
|
La
función coenzimatica de todas las
vitaminas hidrusolubles es razonablemente bien
conocida con excepción de la vitamina
C.se necesitan además otras varias
sustancias hidrosolubles como factores de crecimiento para algunos organismos
pero se encuentran en cantidades mayores que ellas; se incluyen en este grupo
el inositol, la colina y la carnitina.
Las
vitaminas liposulubles comprenden a las
vitaminas A, D, E y K solamente los animales superiores parecen precisarlas de
procedencia exógena; no se ha podido establecer todavía claramente el papel
esencial de las vitaminas liposolubles en los vegetales y en el microorganismo.
No parece que sirva de componentes de
los coenzimas, sin que actúan por otros caminos que precisan solamente
cantidades mínimas de ellas.
La mayoría de las vitaminas hidrosolubles son componentes de
coenzimas.
|
COENZIMAS DERIVADOS DE
ALGUNAS VITAMINAS HIDROSOLUBLES
|
|
|
Vitaminas
|
Coenzimas
derivados
|
|
Tiamina (vitamina B1)
|
Tiamina pirofosfato
|
|
Ribloflavina (vitamina B2
|
Flavina adenina dinucleótido y flavina mononucleótido
|
|
Nicotinato (niacina)
|
Nicotinamida adenina dinucleótido
|
|
Piridoxina, piridoxal y piridoxamina (vitamina B6)
|
Piridoxal fosfato
|
|
Pantotenato
|
Coenzima A
|
|
Biotina
|
Se una covalentemente a las caboxilasas
|
|
Folato
|
Tetrahidrofolato
|
|
Cobalamina
(vitamina B12)
|
Coenzimas
de cobamida
|
LAS VITAMINAS SON PRECURSORES DE
COENZIMAS
El
descubrimiento de que se necesitaban, para la buena nutrición otras sustancias
de además de sales, proteínas, grasas y carbohidratos (sustancias que F. G.
Hopkins llamo factores nutritivos accesorios y Casimir Funk, en 1911, bautizo
con el nombre de vitaminas impulso a los investigadores a buscar la función de
estas sustancias en el metabolismo, para cerciorarse de el motivo de que
debiesen figurar en la alimentación de ciertos organismos. Actualmente se
acepta que estas sustancias son necesarias para el metabolismo de todos los
organismos, bacterias, plantas verdes, hongos y animales. Sin embargo, muchos
seres pueden sintetizar la cantidad necesaria de estas sustancias, los qe no
pueden sintetizarla necesariamente deben obtenerlas con su alimentación. Ya se
conoce la función específica de muchas de estas vitaminas en el metabolismo en
cada caso se sabe que forman parte de una molécula mayor que funciona como
coenzima, la cual, con la enzima y el sustrato, es absolutamente necesaria para
alguna reacción particular. Las enfermedades por carencia (pelagra, escorbuto,
raquitismo y beriberi), debidas a falta de vitaminas, son manifestaciones
típicas del trastornó metabólico que abusado por la coenzima deficiente.
|
ALGUNOS TRANSPORTADORES ACTIVADOS DEL METABOLISMO
| |
|
MOLÉCULA PORTADORA
|
GRUPO TRANSPORTADO EN FORMA ACTIVA
|
|
ATP
|
Fosforilo
|
|
NADH y NADPH
|
Electrones
|
|
FADH2
|
Electrones
|
|
FMNH2
|
Electrones
|
|
Coenzima A
|
Acilos
|
|
Lipoamida
|
Acilos
|
|
Tiamina Pirofosfato
|
Aldehídos
|
|
Biotina
|
CO2
|
|
Tetrahidrofolato
|
Fragmentos monocarbonados
|
|
S-adenosilmetionina
|
Metilo
|
|
Uridina difosfato glucosa
|
Glucosa
|
|
Citidina difosfato glucosa
|
Fosfatidato
|
|
Nucleósido trifosfato
|
Nucleótidos
|
NAD Y NADP
Las coenzimas de nicotinamida
son nicotinamida adenina dinucleótido (NAD) y el fostato de nicotinamida
adenina dinucleótido (NADP), muy relacionado. Fueron ñas primeras coenzimas que
se conocieron. Ambas contienen nicotinamida, la amida del acido nicotínico
(figura 7.7). El ácido nicotínico (llamado también niacina) es el factor que
falta en la pelagra. Es esencial como precursor del NAFD Y NADP. (En muchas especies, el
triptófano se degrada a ácido nicotínico. En consecuencia, el triptófano en la
dieta puede suplir algo de los requisitos de niacina o nicotinamida). Como el
ácido nicotínico es el derivado de piridina con 3-carboxilo, con frecuencia se
llama coenzimas nucleótidos de piridina a las coenzimas de nicotinamida.

Las coenzimas de nicotinamida
participan en muchas reacciones de oxidación- reducción. Ayudan a la
trasferencias de electrones hacia metabolitos y desde estos. Las formas
oxidadas, NAD y NADP, carecen de electrones, y las formas reducidas, NDDH y
NADPH, tienen un par adicional de electrones en forma de ion hidruro unido en
forma covalente. Las estructuras de estas coenzimas se ven en la figura 7.8.
Las dos coenzimas contienen un enlace fosfoanhídrido que une dos
5’-nucleotidos: AMP y el robonucleótido de la nicitinamida, llamado
nicotinamida monucleótido (NMN) (formado
aparte del ácido nicotínico).En el caso del NADP, hay un grupo fosforilo en el
átomo de oxígeno 2’ del adenilato.
En la forma oxidada de la
coenzima (NAD Y NADP), al nitrógeno de la nicotinamida le falta uno de sus
electrones. Sólo tienen cuatro electrones en la capa externa, y están
compartidos con átomos adyacentes de carbono para formar un total de cuatro
enlaces covalentes. (Cada enlace tiene un par de electrones, de modo que la
capa externa del átomo de nitrógeno está llena con ocho electrones
compartidos). Es la causa de que la
carga positiva se asocie normalmente al átomo de nitrógeno del anillo, como se
ve en la figura 7.8. De hecho, la carga está distribuida sobre todo el anillo
aromático.
En particular, el átomo de
nitrógeno tiene cinco electrones en su capa externa. Dos de ellos En la forma
reducida, el anillo aromático tiene su complemento de electrones normal y
completo. (Representados con puntos en la figura 7.8)
Son un par libre de
electrones. Los tres electrones participan en tres enlaces covalentes.
Casi siempre, el NAD y el NADP
actúan como consustratos para deshidrogenasas. Las deshidrogenasas dependientes
de nucleótidos de piridina catalizan la oxidación de sus sustratos,
transfiriendo dos electrones y un proton en forma de un ion hidruro (H) al C-4
del grupo nicotinamida del NADH o NADPH, donde se ha formado un nuevo enlace
C-H en el C-4 (un par de electrones) y
el electrón que antes estaba asociado al doble enlace del anillo se ha
deslocalizado al átomo de nitrógeno del anillo. Así, la oxidación por
nucleótidos de piridina (o la reducción, la reacción inversa) siempre se
efectúa de dos en dos electrones.
Se dice que el NADH y el NADP,
que son estables en soluciones que contienen oxígeno, tiene poder reductor (es
decir, son reductores biológicos). La estabilidad de los nucleótidos de piridina
reducidos les permite llevar su poder reductor de una enzima a otra, propiedad
que no comparten las coenzimas derivadas de riboflavina (sección 7.5). La mayor
parte de las reacciones que forman NADH Y NADPH son catabólicas. La oxidación
de NADH en las mitocondrias esta acoplada a la síntesis del ATP. La mayor parte
del NADPH se usa como agente reductor en reacciones de biosíntesis.
FA D y F MN

Las coenzimas flavina adenina
dinucleótido (FAD) y flavina mononucleótido (FMN) se deriva de la riboflavina o
vitamina B2. La riboflavina es sintetizada por bacterias, protistas,
hongos plantas, y algunos animales. Los mamíferos obtienen riboflavina de su
alimento. La riboflavina está formada por ribitol, un alcohol con cinco
carbonos unidos al átomo N-10 de un sistema de anillo heterociclo llamado
isoaloxazina (figura 7.10a). Las coenzimas derivadas de la riboflavina se ven
en la figura 10b. Igual que el NAD y el NADP, el FAD contiene AMP y un enlace
de pirofosfato.
Las coenzimas de flavina
reducida se pueden oxidar rápidamente en presencia de oxígeno. Como están
firmemente unidas al grupo prostético, las apoenzimas protegen a las formas
reducidas contra la reoxidación, que sería un desperdicio.

COENZIMA A

Muchos procesos metabólicos
dependen de la coenzima A (CoA o HS-CoA), como la oxidación de moléculas de
combustible y la biosíntesis de algunos carbohidratos y lípidos. Esta coenzima
interviene en reacciones de transferencia de grupo acilo, donde los grupos
metabólicos móviles son ácidos carboxílicos y ácidos grasos simples. La
coenzimas A tiene dos componentes principales: una unidad de
2-mercaptoetilamina, que tiene un grupo – SH libre, la vitamina pantotenato-
(vitamina B5, una amida de
B-alanina y pantoato), y una mitad de ADP, cuyo grupo hidroxilo 3’ está
esterificado con un tercer grupo fosfato (figura 7.12a)
COENZIMA DE FLAVINA
El dinucleotido de flavina y
adenina o FAD, es una de las dos coenzimas derivadas de la vitamina B2
o riboflavina. La otra es la más sencilla mononucleotido de flavina (FMN), o
riboflavina fosfato. La parte funcionan de ambas coenzimas es el sistema del
anillo de isoaloxazina, que actúa como aceptor de electrones. Los compuestos
que contienen un anillo de este tipo se denominan flavinas. En la riboflavina y
sus derivados, el sistema de anillo está unido al ribitol, una versión de
cadena abierta de la ribosa, con el carbono aldehído reducido al nivel de
alcohol. El carbono 5’ dl ribitol está ligado al fosfato en el FMN, y el FAD es un derivado adenililado del
FMN. Así pues estos compuestos son análogos en cierta medida al mononucleotido
de nicotinamida y al NAD+ respetivamente.
Las enzimas que utilizan una
coenzima de flavina se denominan flavoproteinas. El FMN y el FAD experimentan
reacciones de transferencia de electrones prácticamente idénticas. Las enzimas
flavoproteicas se unen preferentemente al FMN o al FAD. En unos pocos casos,
esa unión es covalente. Sin embargo, en la mayor parte de los pasos, la flavina
está unida de forma ajustada aunque no covalente, de modo que la coenzima puede
disociarse reversiblemente de la holoenzima.
Como las coenzimas de la
nicotinamida, las flavinas experimentan reacciones de oxidación y reducción de
dos electrones. Sin embargo, las flavinas se diferencian por tener una especie
estable reducida de un electrón, un radical libre de semiquinona.
e
COENZIMAS FOSFORILANTES
Ejemplo:
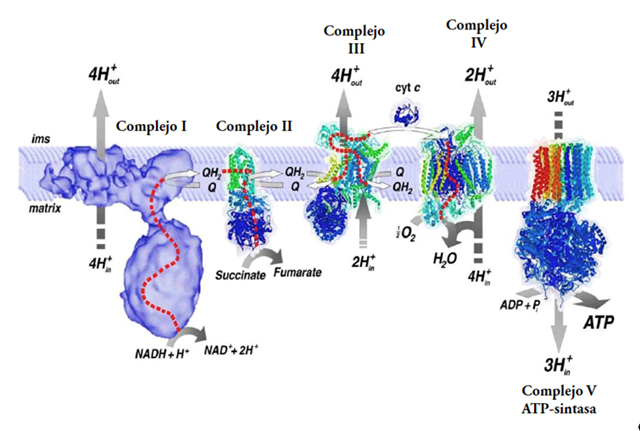
La respiración anaerobia es en
realidad un proceso de desintegración, pero no de glucosa libre, sino de un
derivado fosforilado de la misma. El primer paso en el metabolismo anaerobio
incluye por consiguiente la conversión de la glucosa en su forma éster-fosfato.
La coenzima fosforilante ATP, consiste de la unión química de la adenosina
(adenina más ribosa) y tres fosfatos, la cual sirve en la transformación de la
glucosa a su éster fosfato correspondiente. Reacción que se lleva a cabo por
transferencia enzimática especifica del agrupamiento terminal fosfato del
ATP a glucosa para formar ADP y glucosa
fosfato.2
La glucosa-fosfato es
transformada en enzimáticamente a fructuosa-fosfato, la cual es fosforilada
enzimáticamente por el ATP para formar fructuosa-disfosfato
7.7 Pirofosfato de tiamina
La tiamina (o vitamina B1)
contiene un anillo de pirimidina y un anillo de tiazolio con carga positiva
(figura 7.14). En los mamíferos, la tiamina es una vitamina esencial. Abunda en
las cáscaras de arroz y en otros cereales. La coenzima es el pirofosfato de
tiamina (figura 7.14). También el TPP es una coenzima que interviene en la descarboxilación
oxidante de a-cetoácidos distintos del piruvato. El anillo de tiazolio de la
coenzima contiene al centro reactivo.
FOSFATO DE PIRIDOXAL
La familia de vitamina B6
hidrosoluble consiste en tres
moléculas estrechamente relacionadas que sólo difieren en el estado de
oxidación o aminación en el carbono unido a la posición 4 del anillo de
piridina (figura 16a). La vitamina B6, con mayor frecuencia
piridoxal o piridoxamina, se encuentra con facilidad en muchas fuentes
vegetales y animales. Las deficiencias de B6 inducidas en ratas
causas dermatitis y diversas alteraciones relacionadas con el metabolismo de
las proteínas, pero en realidad son raras las deficiencias de B6 en humanos.
BIOTINA
La biotina es un grupo
protestico para enzimas que catalizan reacciones de transferencia del grupo
carboxilo y reacciones de acrboxilacion dependientes de ATP. La biotina está
unida en forma covalente al sitio activo de su enzima anfitriona por un enlace
de amida al grupo E-amino de un residuo de lisina (figura 7.19).
La reacción de piruvato
carboxilasa demuestra el papel de la biotina como portador del dióxido de
carbono (figura 7.20). En esta reacción dependiente del ATP, el piruvato, el ácido de tres
carbonos, reacciones con bicarbonato y forma el axaloacetato, acido de cuatro
carbonos. Biotina, unida a enzimas, es el portador intermedio del grupo
metabólico carboxilo, que es móvil. Todavía no se conoce el mecanismo con el
que se carboxila la biotina.
TETRAHIDROFOLATO
La estructura de la coenzima
folato, que se llama tetrahidrofolato en forma colectiva, difieren de la
vitamina en dos cosas: son compuestos deducidos (5, 6, 7, 8-tetrahidropterinas)
y están modificadas por adición de residuos de glutamato unidos entre sí
mediante enlaces de ᵧ - glutamilamida (figura 7.21 c). La mitad anionica de
poliglutamilo, en general con cinco a seis residuos de longitud, participa en
el enlazamiento de las coenzimas a las enzimas. Cuando se usa el termino
tetrahidrofolato se debe considerar que se refiere a compuestos que tienen
colas de poliglutomato de diversas longitudes.
Otra coenzima de pterina, la
5, 6, 7, 8-tetrahidrobiopterina, tiene una cadena lateral de tres carbonos en
el C-6 de la mitad de pterina, en vez de la gran cadena lateral que hay en el
tetrahidrofolato (figura 7.23). Esta coenzima no se deriva de alguna vitamina,
si no que los animales y otros organismos la sintetizan.
COBALAMINA
La cobalamina (vitamina B2) es
la mayor de las vitaminas B, y fue la última que se aisló. La estructura de la
cobalamina (figura 7.24a) tiene un sistema corrin anular que se asemeja al
sistema anular de porfirina en el hemo (figura 4.39).
Algunas especies de bacterias
sintetizan la cobalamina. Todos los animales la requieren como micronutrientes,
y también algunas bacterias y algas. Las plantas no requieren cobalamina, por
lo que no la sintetizan. En consecuencia, y en el caso normal, los humanos
obtienen la vitamina B12 a partir de los alimentos de origen animal. Con
frecuencia, los vegetarianos logran cantidades adecuadas producidas por
microorganismos.
LIPOAMIDA
La
coenzima lipoamida es la forma de ácido lipoico unida a proteína. Aunque a
menudo se dice que el ácido lipoico en una vitamina B, parece que los animales
pueden sintetizarlo. Lo requieren ciertas bacterias y protozoarios para crecer.
A
continuacion, el grupo acilo se trasfiere al átomo de azufre de una molécula de
coenzima A para generar la forma reducida (dihidrolipoamina) del grupo
prostético.
Las
acciones de las diversas coenzimas del complejo piruvato deshidrogenasa
muestran como las coenzimas, al proporcionar grupos reactivos que aumentan la
versatilidad catalítica de las proteínas, se usan para conservar tanto la
energia como elementos constructivos carbonados.
VITAMINAS
LIPÍDICAS
Las
estructuras de las cuatro vitaminas lipídicas (A, D, E y K) contienen anillos y
largas cadenas laterales alifáticas. Las vitaminas lipídicas son muy hidrofóbicas,
aunque cada una posee cuando menos un grupo polar.
Vitamina A
La
vitamina A, o retinol, es una molécula lipídica con 20 carbonos, que se obtiene
en la dieta, ya sea en forma directa o indirecta, como β-caroteno. Las
zanahorias y otras verduras amarillas son ricas en β-caroteno, un lípido
vegetal con 40 carbonos cuya ruptura oxidante enzimática produce la vitamina A
(figura 7.27). La vitamina A existe en
tres formas que difieren en estado de oxidación del grupo funcional terminal:
el retinol, un alcohol estable, el retinal, un aldehído, y el ácido retinoico.
Los tres compuestos tienen funciones biológicas importantes.
VITAMINA K
La
vitamina K (filoquinona) (figura 7.29) es una vitamina lipídica procedente de
plantas, necesaria en la síntesis de algunas de las proteínas que intervienen
en la coagulación sanguínea. En una coenzima de una carboxilasa de mamíferos
que cataliza la conversión de residuos específicos de glutomato para formar
residuos de y-carboxiglutamat. La forma reducida (hidroquinona) de la
vitamina K participa en la carboxilacion como agente
reductor.
UBIQUINONA
La
Ubiquinona llamada llamada también coenzima Q y en consecuencia se abrevia Q,
en es una coenzima soluble en lípidos, sintetizada por casi todas las especies.
Es una benzoquinona con cuatro sustituyentes, uno de los cuales es una larga
cadena hidrofóbica (fig. 7.30a). Esta cadena, formada por 6 a 10 unidades por
isoprenoides, permite que la Ubiquinona se disuelva en los lípidos de las
membranas. En la membrana, la Ubiquinona transporta entre complejos enzimáticos
embebidos en la membrana. Algunas bacterias usan menaquinona en vez de
Ubiquinona. Un análogo de la Ubiquinona, llamado plastoquinona (fig. 7.30b)
desempeña una función similar en el transporte fotosintético de cloroplastos
(capitulo 15).
La
Ubiquinona es un agente oxidante más enérgico que NAD o que las coenzimas de la
flavina. En consecuencia, lo pueden reducir NADH o FADH2-. Igual que
el FMN y el FAD, la Ubiquinona puede aceptar o donar dos electrones, uno o dos
al mismo tiempo, porque tiene tres estados de oxidación: Q oxidado, radical
libre de semiquinona parcialmente reducida y totalmente reducida QH2-,
llamada ubiquinol (fig. 31). La coenzima Q tiene un papel principal en el
transporte de electrones asociado a la membrana. Es responsable de mover
protones de uno a otro lado de la membrana, por un proceso llamado ciclo Q
(capitulo 14). El gradiente de protones que resulta impulsa la síntesis de ATP.
Proteínas
coenzimas
Algunas
proteínas funcionan como coenzimas. No catalizan reaccionan ellas mismas, pero
ciertas enzimas la necesitan. Estas coenzimas se llaman proteínas de
transferencia de grupo, o proteínas coenzimas.
Contiene un grupo funcional que es parte de la columna vertebral de la
proteína, o bien es un grupo prostético. En general son más pequeñas y más termoestables
que la mayor parte de las enzimas. Las proteínas coenzimas se llaman coenzimas
por que participan en muchas y diversas reacciones, y se asocian a una variedad
de enzimas diferentes.
Algunas
proteínas coenzimas participan en reacciones de transferencia de grupo, o en
reacciones de oxidación-reducción donde el grupo transferido es Hidrogeno o un
electrón. Los iones metálicos, grupos de hierro-azufre y grupos hemo son
centros reactivos que suelen encontrar en estas coenzimas.
CITOCROMOS
Los
citocromos son proteínas coenzimas que contienen hemo, cuyos átomos de Fe (III)
sufren reducción reversible de electrón. En las figuras 4.21 y 4.24b se
mostraron algunas estructuras de citocromos. Los citocromos pueden ser de clase
a, b y c, de acuerdo con sus espectros de absorción visible. Las clases tienen
grupos hemo prostéticos un poco diferentes (figura 7.33). El grupo hemo de los
citocromos tipo b es igual al de hemoglobina y mioglobina (figura 4.44). El
hemo del citocromo a tiene una cadena hidrofóbica de 17 carbonos en el C-2 del
anillo de porfirina, y un grupo formilo en el C-8, en tanto que el hemo tipo b
tiene un grupo vinilo unido al C-2 y un grupo metilo en C-8. En los citocromos
tipo c, el hemo está unido en forma covalente a la aproteina por grupos vinilo
del hemo mediante dos enlaces tioéter que se forman por adición los grupos tiol
de dos residuos de cisteína.
UTILIZACIÓN DE LOS NUCLEÓTIDOS DE
ADENINA EN LA BIOSÍNTESIS DE COENZIMAS
Una función
metabólica importante de los nucleótidos de purina es la que desempeñan en la
síntesis de coenzimas, fundamentalmente las que contienen un grupo adenilato.
Entre ellas se encuentran los nucleótidos de flavina, los nucleótidos de
nicotinamida y la coenzima A, como se muestra en el siguiente resumen:

Bibliografía
Lehninger, A. L. (1995). BIOQUIMICA
(las bases moluculares de la estructura y funcion celular). Barcelona :
OMEGA.
STRYER, L. (2001). BIOQUÍMICA
. Barcelona : cuarta .
Bioquímica, P. d. (2008). H. Robert Horton, Laurence
A. Moran, K. Gray Scrimgeour, Marc D. Perry, J. David Rawn. MEXICO :
cuarta edicion
MATHEWS, C., VAN HOLDE, K., & AHERN, K. (2002). BIOQUIMICA.
Madrid: Pearson.
Nason, A. (2002). Biologia . Mexico, D.F.: Limusa.
VILLEE, C. A. (1996). BIOLOGIA. Mexico, D.F.: McGraw-hill.
No hay comentarios.:
Publicar un comentario